





Dr Nathaniel T. Jeanson
Answers in Genesis
W dyskusji o powstawaniu gatunków jedną z kluczowych kwestii jest dryf genetyczny. Mało kto o nim słyszał, ale pojęcie to odgrywa szczególnie istotną rolę w odwracaniu do góry nogami współcześnie dominującego naukowego wyjaśnienia powstawania gatunków – oraz całej debaty ewolucjonizm-kreacjonizm.
Tło
Przez lata pojęcie dryfu genetycznego pozostawało w cieniu o wiele bardziej znanej koncepcji – doboru naturalnego lub inaczej przetrwania najlepiej przystosowanych do wydania potomstwa. Łatwo zauważyć, dlaczego tak było. W końcu Darwin użył tego terminu w tytule swojej książki: „O powstawaniu gatunków drogą doboru naturalnego”. Do dzisiaj w wyjaśnianiu projektu obserwowanego w przyrodzie ewolucjoniści mocno opierają się na koncepcji doboru naturalnego, a nie dryfu genetycznego.
[…] w przeciwieństwie do doboru naturalnego dryf genetyczny nie wytwarza adaptacji, dlatego ten proces nie warunkuje tych cech anatomicznych, fizjologicznych i behawioralnych, które pozwalają organizmom na przeżycie i rozród. 1
Drugim czynnikiem, który na 150 lat zapewnił dominację idei doboru naturalnego w debacie o powstawaniu gatunków, jest prostota tej koncepcji – lepsi przetrwają, gorsi wymrą.
Dryf genetyczny pozostał natomiast pojęciem niejasnym i obcym dla większości społeczeństwa. W skrócie można powiedzieć, że chodzi w nim o zastosowanie statystyki do genetyki. Proszę się jednak nie obawiać: wykorzystywana tu matematyka jest dość łatwa do zrozumienia – i w zdumiewający sposób odnosi się do kwestii powstawania gatunków. Wkrótce przekonamy się, że dryf genetyczny odgrywał prawdopodobnie jedną z głównych ról w powstawaniu gatunków z przedstawicieli „rodzajów” [ang. kinds] zwierząt zabranych do arki Noego. 2
Nowe gatunki i potop
Kiedy zajmujemy się dryfem genetycznym, warto przede wszystkim rozumieć, że nie mamy do czynienia z prawami nauki, ale z prawdopodobieństwami. Przykładowo, jeśli wziąć pod uwagę zbiór wszystkich przyszłych rodziców, jakie prawo nauki pozwala przewidzieć, czy będą mieli syna, czy córkę? Mimo że żyjemy w zaawansowanych pod względem naukowym czasach, nie dysponujemy tego rodzaju prawem – dlatego imprezy z okazji ujawnienia płci dziecka są tak popularne wśród rodziców spodziewających się potomstwa! 3 Jako że nie ma odpowiedniego prawa, uciekamy się do prawdopodobieństwa. Średnio wśród urodzonych dzieci około 50% to chłopcy i 50% to dziewczynki. Występują odchylenia od tej średniej – dlatego właśnie mówimy o prawdopodobieństwach, a nie o prawie. Ze statystycznego punktu widzenia większość rodziców ma 50% szans, że urodzi im się chłopiec i 50% szans, że urodzi się dziewczynka. W niniejszym tekście zbadamy kwestię powstawania gatunków w ujęciu probabilistycznym.
Rozważmy na przykład sprawę powstania siedmiu żyjących gatunków z rodziny koniowatych (tj. konie, osły i zebry; patrz ryc. 1). Na podstawie badań dotyczących możliwości ich krzyżowania sądzimy, że należą one zapewne do tego samego biblijnego rodzaju, a w związku z tym – że pochodzą od jednej pary koniowatych, które weszły do arki. Para tych zwierząt po potopie opuściła arkę i przez następne 4500 lat z ich potomstwa wyłoniło się siedem żyjących obecnie gatunków. Rozważmy teraz bardziej szczegółowo, jaką rolę w tym procesie mógł odegrać dryf genetyczny.

Przyjmijmy na początek, że Noe zabrał do arki heterozygotyczną parę rodzaju koniowatych – czyli parę, która nie miała takiego samego DNA u obojga przedstawicieli, ale miała wiele różnic w informacji genetycznej (dla przypomnienia: przedrostek homo- znaczy „taki sam”, a hetero- znaczy „różny”). U stworzeń rozmnażających się płciowo potomstwo dziedziczy instrukcje genetyczne od obojga rodziców. Źrebię zebry połowę swojego DNA otrzymuje od matki, a drugą połowę – od ojca. Ale nie jest tak, że od ojca pochodzi informacja genetyczna dotycząca prawej części ciała, a od matki – dotycząca lewej części. Każde z rodziców przekazuje informację dotyczącą całego ciała, a u potomka wykorzystywana jest pewna kombinacja tej informacji i w ten sposób powstaje jego niepowtarzalny wygląd – dzieje się tak dzięki heterozygotycznej informacji genetycznej uzyskanej przez dziedziczenie od dwojga różniących się rodziców.
Dla jasności, nie chodzi o to, że DNA samca z pary zwierząt koniowatych było całkowicie inne niż DNA samicy. Aktualne dane wskazują, że w ogromnej większości sekwencje DNA dwu osobników były jednakowe. 4 Jednak sekwencje te różniły się w około 25 milionach (lub więcej) miejsc (wbrew temu, co mogłoby się wydawać, jest to tylko mała część całego DNA!).
Jak ta jedna para mogła dać początek zarówno mającym, jak i niemającym pasków gatunkom koni, które dzisiaj chodzą po Ziemi?
Dryf genetyczny i potop
Aby ułatwić śledzenie dalszych rozważań, uprościmy nieco genetykę. Przyjmijmy, że A będzie oznaczało informację genetyczną odpowiedzialną za powstawanie pasków, natomiast a – informację dla jednolitego umaszczenia. (W naszym przykładzie zakładamy, że spośród 25 milionów miejsc sekwencji DNA, w których występują różnice, tylko jedno miejsce ma znaczenie w kwestii umaszczenia – i to również jest uproszczeniem). Heterozygotyczna para zwierząt, która weszła do arki, w części DNA decydującej o kolorze sierści miałaby informację genetyczną Aa – zarówno samiec, jak i samica – każde z nich byłoby heterozygotą Aa, czyli miałoby oba warianty instrukcji genetycznej, tj. A i a. Zwierzęta te byłyby częściowo w paski, podobnie jak hybrydy konia i zebry oraz osła i zebry, które można zobaczyć w zoo przy Muzeum Stworzenia (Creation Museum) w Stanach Zjednoczonych.

Wykorzystując tzw. szachownicę Punnetta, możemy przewidzieć, jakie jest prawdopodobieństwo wydania przez tę parę rodzicielską potomstwa, które miałoby paski lub jednolite umaszczenie.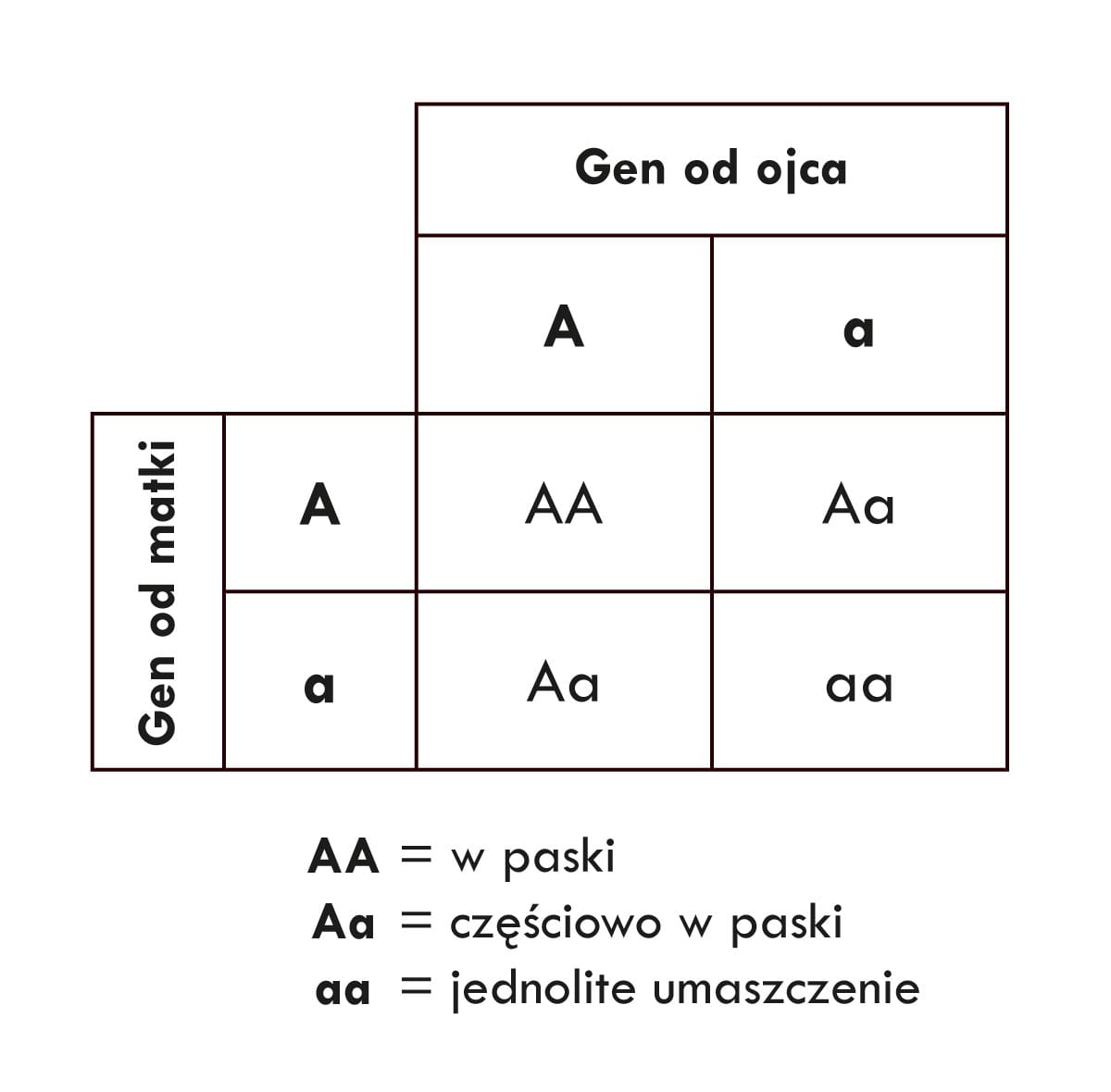
Średnio więc około 25% potomstwa (1 na 4) będzie miało paski (te z informacją genetyczną AA), 25% (1 na 4) będzie miało jednolite umaszczenie (czyli kombinację genów aa), a 50% będzie częściowo w paski, tak jak ich rodzice (kombinacja genów Aa). Inaczej mówiąc, potomstwo w tym wypadku ma 25% szans, że będzie miało sierść w paski, 25% szans na jednolite umaszczenie i 50% szans, że będzie podobne do rodziców (czyli częściowo w paski).
Jaki to ma związek z pytaniem o specjację, czyli o powstawanie gatunków? Nie będziemy tu przyjmować znanego, choć zapewne błędnego założenia, że specjacja prowadzi do określonego celu. Proces powstawania gatunków w środowisku naturalnym nie jest tym samym, co proces hodowli różnych ras na farmach i ranczach. Aby powstał nowy gatunek, potomstwo nie musi spełnić jakiegoś wcześniej ustalonego celu czy dążenia. Do tego, żeby dana grupa zwierząt została uznana za nowy gatunek, nie musi ona mieć jakiejś określonej cechy, którą akurat chcieliby w niej widzieć biolodzy. Populacje zwierząt uzyskują miano nowych gatunków, jeśli grupy te odróżniają się i są odizolowane od pozostałych populacji. Dla celów naszej dyskusji chcemy więc wiedzieć, jakie jest prawdopodobieństwo, że potomstwo będzie w jakiś sposób odróżniało się wyglądem od rodziców.
W naszym przykładzie od swoich rodziców różnią się potomkowie zarówno o jednolitym umaszczeniu, jak i w paski. Prawdopodobieństwa tych dwóch przypadków stanowią łącznie prawdopodobieństwo pojawienia się potomstwa odróżniającego się od rodziców: 25% + 25% = 50% szans. Nieźle!
Zatrzymajmy się chwilę i pomyślmy o tym, zanim przejdziemy do rozważań nad obrazem statystycznym kolejnego pokolenia. Wśród potomstwa, które pojawia się w pierwszym pokoleniu, nie powstał jeszcze nowy gatunek. Do tego potrzebne byłoby utworzenie całej populacji osobników, które wyglądałyby podobnie do siebie nawzajem, ale różniłyby się od rodziców. Średnio 5 u koniowatych rodzi się rocznie jedno młode, które osiąga dojrzałość płciową dopiero po około 3 latach (ze względu na multiplikatywny charakter wzrostu, nawet przy tym początkowo powolnym tempie powiększania się populacji może ona osiągnąć ponad milion osobników w ciągu stu lat! 6). Aby doszło do powstania nowego gatunku, musi się pojawić w naszych statystykach dwoje potomków różniących się wyglądem od rodziców.
Mówiąc dokładniej, potrzebnych jest dwoje potomków różnych od swoich rodziców, ale podobnych do siebie nawzajem. Gdyby jedno z nich było w paski (czyli AA), a drugie jednolicie umaszczone (czyli aa), to ich potomstwo po prostu wyglądałoby znowu tak samo jak rodzice rodziców – byłoby częściowo w paski (czyli Aa), nie zaszlibyśmy więc za daleko, jeśli chodzi o powstanie nowego gatunku.
Innymi słowy, potrzebnych jest dwoje potomków w paski lub dwoje potomków z jednolitym umaszczeniem. Prawdopodobieństwo takiego rezultatu jest równe szansom uzyskania jego dwóch składowych pod rząd – czyli 25% (1 na 4) pomnożone przez 25% (1 na 4), co daje prawdopodobieństwo wynoszące 1 na 16. Zamierzony rezultat może być jednak osiągnięty na dwa sposoby, z których każdy osobno ma 1 na 16 szans wystąpienia – albo dwoje potomków w paski (szanse 1 na 16), albo dwoje potomków jednolicie umaszczonych (szanse również 1 na 16). Łącznie daje to szanse wynoszące 2 na 16 (1 na 16 + 1 na 16). Zatem z punktu widzenia statystyki para zwierząt z rodzaju koniowatych, która przetrwała w arce, mogła wydać na świat dwoje potomków różnych od rodziców, ale podobnych do siebie nawzajem z prawdopodobieństwem tylko 12,5%. Inaczej mówiąc, w 87,5% przypadków nie udałoby się parze rodziców wydać na świat pary potomków, którzy mogliby dać początek nowemu gatunkowi. Czy w związku z tym powstawanie gatunków po potopie było czymś trudnym?
Nie. Tutaj właśnie odgrywa rolę wielopokoleniowy statystyczny efekt dryfu genetycznego. Powiedzmy na przykład, że z pary koni, które były w arce Noego, urodziło się najpierw młode w paski (AA), a następnie młode częściowo w paski, tak jak rodzice (Aa). Dla scenariusza, w którym pojawia się jeden potomek homozygotyczny – czyli AA lub aa – i jeden potomek heterozygotyczny – czyli Aa – prawdopodobieństwo wynosi 50%. 7 Jeśli z kolei ta para potomków będzie miała własne młode, prawdopodobieństwa dla nich możemy obliczyć za pomocą innej szachownicy Punnetta: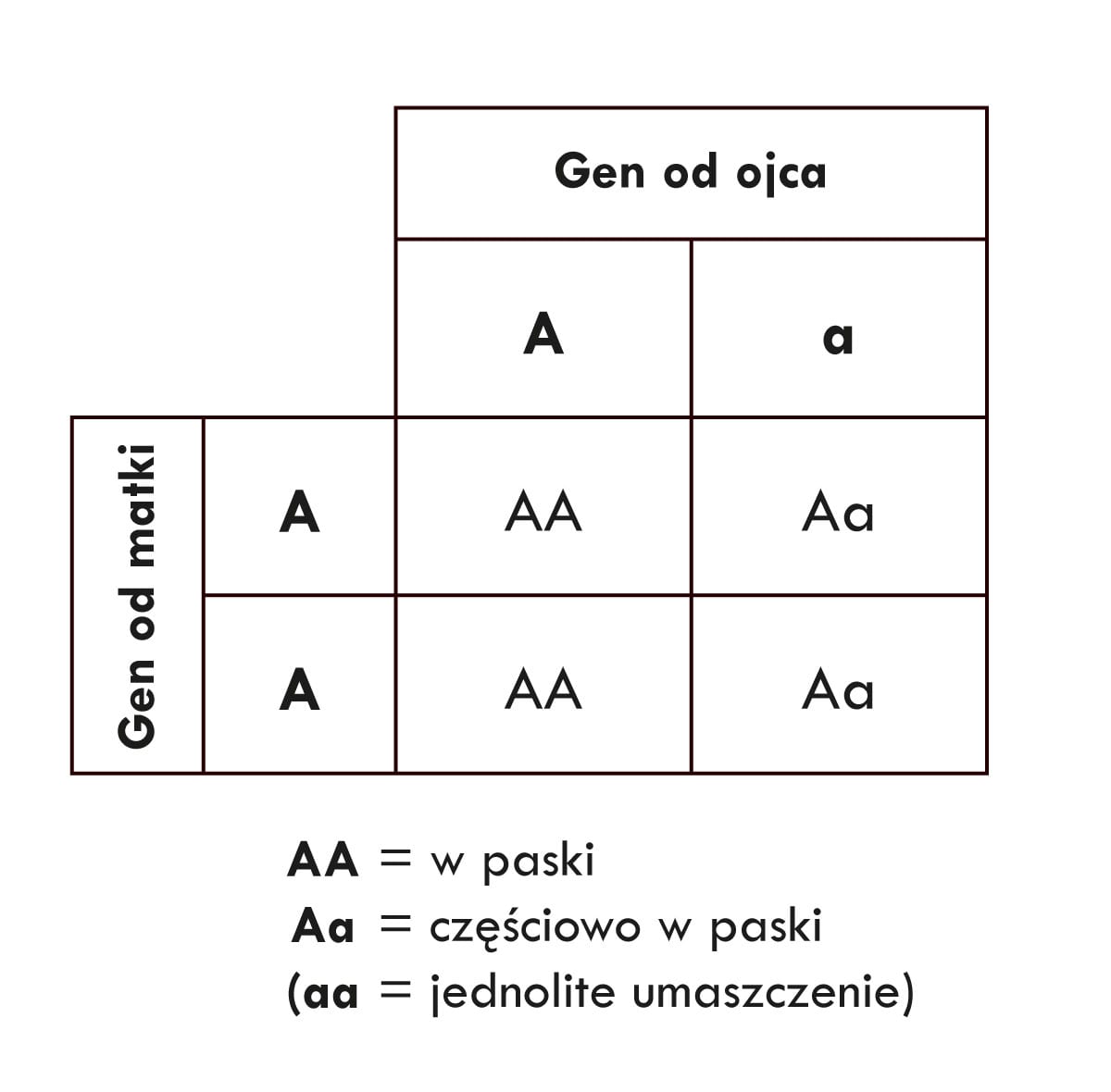
Tutaj także szanse na to, że młode będą wyglądały inaczej niż para rodzicielska z arki (w tym wypadku szanse pojawienia się potomka z układem genów AA), wynoszą 50% (tj. 1 na 2). Wzrastają jednak szanse pojawienia się po sobie dwójki młodych podobnych do siebie, ale różniących się od przodków z arki. Tak samo jak poprzednio, mnożymy prawdopodobieństwa dla każdego z tych potomków – czyli prawdopodobieństwo uzyskania AA mnożymy przez prawdopodobieństwo uzyskania AA. Obliczenia te pokazują, że w przypadku krzyżowania się pary AA i Aa, szanse na pojawienie się dwójki młodych, które mogą dać początek nowemu gatunkowi, wynoszą 25% (1 na 2 mnożymy razy 1 na 2, co daje 1 na 4, czyli 0,25 lub inaczej 25%).
Zatem w ciągu zaledwie dwóch pokoleń szanse pojawienia się wśród potomstwa pary, która może dać początek nowemu gatunkowi, wzrastają do 1/4. Każdy uczestnik loterii byłby zachwycony takim prawdopodobieństwem wygranej. Gdyby zaś populacja ta została odizolowana od pozostałych przedstawicieli swojego rodzaju – co bez trudu mogło nastąpić, kiedy zwierzęta rozeszły się na całą ziemię spod góry Ararat – stałaby się populacją homozygotyczną. Inaczej mówiąc, w tej grupie zwierząt zostałaby utracona część informacji genetycznej, która była obecna u pary rodzicielskiej – informacja A lub, jak w naszym przykładzie, a. W efekcie doprowadziłoby to do trwałej zmiany w populacji, którą należałoby wówczas uznać za nowy gatunek.
Jednak czy nie ma tu znaczenia fakt, że uprościliśmy omawiany proces? W przypadku bardziej realistycznego scenariusza sprawa byłaby jeszcze łatwiejsza. Większa liczba różnic w DNA oznacza więcej możliwości powstania potomstwa odróżniającego się od rodziców – co daje więcej możliwości powstania nowego gatunku!
Szybkie powstawanie gatunków
Rozważmy to, co właśnie zaobserwowaliśmy: Jeśli na wstępie mamy heterozygotyczną parę rodziców (a potwierdza to wiele świadectw 8), to szybkie powstanie nowego gatunku jest praktycznie gwarantowane. Trudno zauważyć ten fakt na pierwszy rzut oka. Zapewne dlatego, że składa się na niego kilka z pozoru mało znaczących kroków statystycznych. Jednak małe zmiany, które są możliwe na kolejnych etapach procesu, szybko się kumulują, prowadząc do kompleksowej zmiany, którą nazywamy powstaniem nowego gatunku.
W pewnym sensie omawiany tu scenariusz przypomina przechodzenie po wąskiej grani w górach. Odpowiednikiem grani niech będzie heterozygotyczny stan genetyczny przodków z arki Noego. Powiedzmy, że doliny po obu stronach odpowiadają homozygotycznemu stanowi genetycznemu ich potomków, którzy dali początek nowym gatunkom. Kiedy idziesz wąską granią, od czasu do czasu zdarza się, że przypadkowo postawisz stopę poza wyznaczoną ścieżką. Na tak wąskiej drodze trzeba będzie starać się utrzymać i odzyskać równowagę, żeby nie spaść w dół zbocza. Każde odbicie ze ścieżki wymaga dodatkowego wysiłku, żeby znaleźć się z powrotem na grani. Może się też zdarzyć, że w następnym kroku nadal będziesz poza wytyczonym szlakiem. Każdy krok postawiony poza ścieżką zwiększa ryzyko, że wypadniesz z trasy na stałe – spadając w dół zbocza. Sprawy mają się podobnie we wspomnianych scenariuszach dotyczących genetycznej ścieżki. Trudno utrzymać się na grani. Łatwo zaś można zejść ze szlaku i stoczyć się w dolinę.

Nasza analogia ujawnia też, dlaczego ewolucjoniści o wiele bardziej podkreślają rolę doboru naturalnego niż dryfu genetycznego. Oni także używają podobnej ilustracji, przyrównując miejsca w dolinach do nowych gatunków, a grzbiety gór do stanu heterozygotycznego. Jednak w ich scenariuszu zwierzęta nie zaczynają swojej drogi na grani, ale w dolinach. Wchodzenie z dna doliny na wierzchołek grani – aby stamtąd dostać się do następnej doliny – wymaga nie lada wysiłku i przejścia wielu kroków. Prawdopodobieństwo takiego osiągnięcia jest bardzo małe. Aby więc zwiększyć szanse na to, że tak nieprawdopodobne zdarzenia w końcu się zrealizują, ewolucjoniści odwołują się do ogromnej ilości czasu. Przede wszystkim zaś, starając się „ułatwić” zwierzętom drogę na szczyt góry, odwołują się oni do doboru naturalnego. Dobrze wiedzą, że łut szczęścia nie daje zbyt wielkiej nadziei na przejście przez grań. Liczą jednak na to, że dobór naturalny umacnia przypadkowe małe kroczki w górę, które zdarzają się czasem, gdy zwierzę przemierza dolinę. Jeśli dobór naturalny może utrwalać każdy taki przypadkowy kroczek, to przypuszczalnie zwierzę w końcu jest w stanie znaleźć się na samym wierzchołku góry.
W teorii wyjaśnienie to może się wydawać wiarygodne. W praktyce jednak niektóre ewolucyjne ścieżki obejmują nie tylko doliny i granie, ale także przepaście. Pewne hipotetyczne formy pośrednie znajdują się na ścieżce ewolucyjnej w dolinach oddzielonych od grani głębokim kanionem, którego nie sposób przejść małymi kroczkami wspomaganymi doborem naturalnym. W rzeczywistości – jak stwierdziłby każdy obserwator, patrząc na taką przepaść – tylko jakiś cudowny skok mógłby przenieść zwierzę z doliny na drugą stronę kanionu, skąd potencjalnie mogłoby ono rozpocząć wspinaczkę ku grani. Ponieważ jednak teoria ewolucji wyklucza cudowne skoki, to istnienie głębokich przepaści wskazuje, że pewne ewolucyjne przejścia są niemożliwe. 9
Podsumowując, jednym z kluczy do zrozumienia, jak powstają gatunki, jest dryf genetyczny – proste zastosowanie statystyki do podstawowej genetyki. Jeśli Noe wziął do arki heterozygotycznych przodków, to utworzenie się nowych gatunków – w szybkim tempie – było praktycznie zagwarantowane. A skoro tak, to kreacjoniści prowadzą, gdy chodzi o najważniejszą kwestię w sporze ewolucjonizm-kreacjonizm.
Tekst oryginalny: Nathaniel T. Jeanson, „Secrets to the Origin of Species: Demystifying «Genetic Drift»”, Answers in Genesis, August 16, 2019.
Tłumaczenie: Małgorzata Gazda
Wszystkie prawa zastrzeżone: Copyright © Answers in Genesis. All Rights Reserved. Translated and published by permission of Answers in Genesis. (Answers® and Answers in Genesis® are registered trademarks of Answers in Genesis, Inc.)
Więcej informacji na temat Answers in Genesis znajdziesz na: www.AnswersinGenesis.org, www.CreationMuseum.org i www.ArkEncounter.com.
Przypisy:
1 Douglas J. Futuyma, Ewolucja, Wydawnictwa Uniwersytetu Warszawskiego, Warszawa 2008, s. 228.
2 Por. Nathaniel T. Jeanson, „Jakie zwierzęta zostały zabrane na arkę Noego? Odkrywanie przeszłości”, idzpodprad.pl, 23.11.2020.
3 W krajach anglojęzycznych taką imprezę nazywa się gender-reveal party (przyp. tłum.).
4 Por. Nathaniel T. Jeanson and Jason Lisle, „On the Origin of Eukaryotic Species’ Genotypic and Phenotypic Diversity”, Answers Research Journal 2016, vol. 9, s. 81-122. Por. też Nathaniel T. Jeanson, Replacing Darwin: The New Origin of Species, Master Books, Green Forest 2017.
5 Por. Jeanson, Lisle, „On the Origin of Eukaryotic Species…”, dodatek, tablica 8.
6 Por. j.w.
7 Ze względu na to, że obliczenia są tu dość skomplikowane, nie przedstawiam w tym miejscu pełnego matematycznego uzasadnienia.
8 Por. Jeanson, Replacing Darwin: The New Origin of Species…
9 Więcej na temat tej przeszkody dla ewolucjonizmu można znaleźć w części „Irreducible Complexity” („nieredukowalna złożoność”) artykułu: Kevin Anderson, Brian Catalucci and Nathaniel Jeanson, „Three Puzzles Evolution Can’t Solve”, Answers Magazine 2015, vol. 10, nr 3, s. 58–66.





